


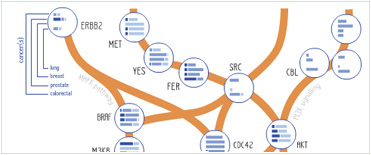







 Foundational characteristics of cancer include proliferation, angiogenesis, migration, evasion of apoptosis, and cellular immortality. Find key markers for these cellular processes and antibodies to detect them.
Foundational characteristics of cancer include proliferation, angiogenesis, migration, evasion of apoptosis, and cellular immortality. Find key markers for these cellular processes and antibodies to detect them. The SUMOplot™ Analysis Program predicts and scores sumoylation sites in your protein. SUMOylation is a post-translational modification involved in various cellular processes, such as nuclear-cytosolic transport, transcriptional regulation, apoptosis, protein stability, response to stress, and progression through the cell cycle.
The SUMOplot™ Analysis Program predicts and scores sumoylation sites in your protein. SUMOylation is a post-translational modification involved in various cellular processes, such as nuclear-cytosolic transport, transcriptional regulation, apoptosis, protein stability, response to stress, and progression through the cell cycle. The Autophagy Receptor Motif Plotter predicts and scores autophagy receptor binding sites in your protein. Identifying proteins connected to this pathway is critical to understanding the role of autophagy in physiological as well as pathological processes such as development, differentiation, neurodegenerative diseases, stress, infection, and cancer.
The Autophagy Receptor Motif Plotter predicts and scores autophagy receptor binding sites in your protein. Identifying proteins connected to this pathway is critical to understanding the role of autophagy in physiological as well as pathological processes such as development, differentiation, neurodegenerative diseases, stress, infection, and cancer.Abcepta Products
CROWN ANTIBODIES
Crown antibodies pass additional stringent quality requirements, including extended control sets, uniform results against multiple biologically relevant cell lines and tissues, and function in multiple applications.
CELL / TISSUES / LYSATES
Abcepta offers a variety of tissue extracts, whole cell lysates and over-expression lysates. View our collections below:
FLcDNA CLONES
Researchers worldwide depend on Abcepta FLcDNA clones to accelerate their research. Expression-ready clones are convenient solution that decreases the time required to start an experiment and increases the chances of success.
NEW PRODUCTS
Abcepta develops 1000+ new products per year. We provide tools for path-breaking research by developing antibodies that detect a comprehensive library of novel and established targets.

Submit your citation using an Abcepta antibody to info@abcepta.com, and receive a free "I Love Antibodies" mug.