


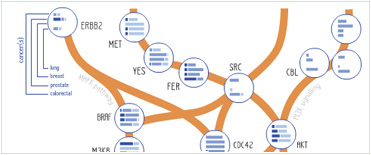







 Foundational characteristics of cancer include proliferation, angiogenesis, migration, evasion of apoptosis, and cellular immortality. Find key markers for these cellular processes and antibodies to detect them.
Foundational characteristics of cancer include proliferation, angiogenesis, migration, evasion of apoptosis, and cellular immortality. Find key markers for these cellular processes and antibodies to detect them. The SUMOplot™ Analysis Program predicts and scores sumoylation sites in your protein. SUMOylation is a post-translational modification involved in various cellular processes, such as nuclear-cytosolic transport, transcriptional regulation, apoptosis, protein stability, response to stress, and progression through the cell cycle.
The SUMOplot™ Analysis Program predicts and scores sumoylation sites in your protein. SUMOylation is a post-translational modification involved in various cellular processes, such as nuclear-cytosolic transport, transcriptional regulation, apoptosis, protein stability, response to stress, and progression through the cell cycle. The Autophagy Receptor Motif Plotter predicts and scores autophagy receptor binding sites in your protein. Identifying proteins connected to this pathway is critical to understanding the role of autophagy in physiological as well as pathological processes such as development, differentiation, neurodegenerative diseases, stress, infection, and cancer.
The Autophagy Receptor Motif Plotter predicts and scores autophagy receptor binding sites in your protein. Identifying proteins connected to this pathway is critical to understanding the role of autophagy in physiological as well as pathological processes such as development, differentiation, neurodegenerative diseases, stress, infection, and cancer.
CUL3 Blocking Peptide (C-term)
Synthetic peptide
- SPECIFICATION
- CITATIONS
- PROTOCOLS
- BACKGROUND
| Primary Accession | Q13618 |
|---|---|
| Other Accession | B5DF89, Q9JLV5, Q6GPF3, Q6DE95, NP_003581.1 |
| Gene ID | 8452 |
|---|---|
| Other Names | Cullin-3, CUL-3, CUL3, KIAA0617 |
| Target/Specificity | The synthetic peptide sequence is selected from aa 751-764 of HUMAN CUL3 |
| Format | Peptides are lyophilized in a solid powder format. Peptides can be reconstituted in solution using the appropriate buffer as needed. |
| Storage | Maintain refrigerated at 2-8°C for up to 6 months. For long term storage store at -20°C. |
| Precautions | This product is for research use only. Not for use in diagnostic or therapeutic procedures. |
| Name | CUL3 (HGNC:2553) |
|---|---|
| Synonyms | KIAA0617 |
| Function | Core component of multiple cullin-RING-based BCR (BTB-CUL3- RBX1) E3 ubiquitin-protein ligase complexes which mediate the ubiquitination and subsequent proteasomal degradation of target proteins. BCR complexes and ARIH1 collaborate in tandem to mediate ubiquitination of target proteins (PubMed:27565346). As a scaffold protein may contribute to catalysis through positioning of the substrate and the ubiquitin-conjugating enzyme. The E3 ubiquitin- protein ligase activity of the complex is dependent on the neddylation of the cullin subunit and is inhibited by the association of the deneddylated cullin subunit with TIP120A/CAND1. The functional specificity of the BCR complex depends on the BTB domain-containing protein as the substrate recognition component. BCR(KLHL42) is involved in ubiquitination of KATNA1. BCR(SPOP) is involved in ubiquitination of BMI1/PCGF4, BRMS1, MACROH2A1 and DAXX, GLI2 and GLI3. Can also form a cullin-RING-based BCR (BTB-CUL3-RBX1) E3 ubiquitin-protein ligase complex containing homodimeric SPOPL or the heterodimer formed by SPOP and SPOPL; these complexes have lower ubiquitin ligase activity. BCR(KLHL9-KLHL13) controls the dynamic behavior of AURKB on mitotic chromosomes and thereby coordinates faithful mitotic progression and completion of cytokinesis. BCR(KLHL12) is involved in ER-Golgi transport by regulating the size of COPII coats, thereby playing a key role in collagen export, which is required for embryonic stem (ES) cells division: BCR(KLHL12) acts by mediating monoubiquitination of SEC31 (SEC31A or SEC31B) (PubMed:22358839, PubMed:27716508). BCR(KLHL3) acts as a regulator of ion transport in the distal nephron; by mediating ubiquitination of WNK4 (PubMed:23387299, PubMed:23453970, PubMed:23576762). The BCR(KLHL20) E3 ubiquitin ligase complex is involved in interferon response and anterograde Golgi to endosome transport: it mediates both ubiquitination leading to degradation and 'Lys-33'-linked ubiquitination (PubMed:20389280, PubMed:21670212, PubMed:21840486, PubMed:24768539). The BCR(KLHL21) E3 ubiquitin ligase complex regulates localization of the chromosomal passenger complex (CPC) from chromosomes to the spindle midzone in anaphase and mediates the ubiquitination of AURKB (PubMed:19995937). The BCR(KLHL22) ubiquitin ligase complex mediates monoubiquitination of PLK1, leading to PLK1 dissociation from phosphoreceptor proteins and subsequent removal from kinetochores, allowing silencing of the spindle assembly checkpoint (SAC) and chromosome segregation (PubMed:23455478). The BCR(KLHL22) ubiquitin ligase complex is also responsible for the amino acid-stimulated 'Lys-48' polyubiquitination and proteasomal degradation of DEPDC5. Through the degradation of DEPDC5, releases the GATOR1 complex-mediated inhibition of the TORC1 pathway (PubMed:29769719). The BCR(KLHL25) ubiquitin ligase complex is involved in translational homeostasis by mediating ubiquitination and subsequent degradation of hypophosphorylated EIF4EBP1 (4E-BP1) (PubMed:22578813). The BCR(KLHL25) ubiquitin ligase complex is also involved in lipid synthesis by mediating ubiquitination and degradation of ACLY (PubMed:27664236). The BCR(KBTBD8) complex acts by mediating monoubiquitination of NOLC1 and TCOF1, leading to remodel the translational program of differentiating cells in favor of neural crest specification (PubMed:26399832). Involved in ubiquitination of cyclin E and of cyclin D1 (in vitro) thus involved in regulation of G1/S transition. Involved in the ubiquitination of KEAP1, ENC1 and KLHL41 (PubMed:15983046). In concert with ATF2 and RBX1, promotes degradation of KAT5 thereby attenuating its ability to acetylate and activate ATM. The BCR(KCTD17) E3 ubiquitin ligase complex mediates ubiquitination and degradation of TCHP, a down- regulator of cilium assembly, thereby inducing ciliogenesis (PubMed:25270598). The BCR(KLHL24) E3 ubiquitin ligase complex mediates ubiquitination of KRT14, controls KRT14 levels during keratinocytes differentiation, and is essential for skin integrity (PubMed:27798626). The BCR(KLHL18) E3 ubiquitin ligase complex mediates the ubiquitination of AURKA leading to its activation at the centrosome which is required for initiating mitotic entry (PubMed:23213400). The BCR(KEAP1) E3 ubiquitin ligase complex acts as a key sensor of oxidative and electrophilic stress by mediating ubiquitination and degradation of NFE2L2/NRF2, a transcription factor regulating expression of many cytoprotective genes (PubMed:15601839, PubMed:16006525). As part of the CUL3(KBTBD6/7) E3 ubiquitin ligase complex functions mediates 'Lys-48' ubiquitination and proteasomal degradation of TIAM1 (PubMed:25684205). By controlling the ubiquitination of that RAC1 guanine exchange factors (GEF), regulates RAC1 signal transduction and downstream biological processes including the organization of the cytoskeleton, cell migration and cell proliferation (PubMed:25684205). The BCR(KBTBD4) E3 ubiquitin ligase complex targets CoREST corepressor complex components RCOR1, KDM1A/LSD1 and HDAC2 for proteasomal degradation with RCOR1 likely to be the primary target while degradation of KDM1A and HDAC2 is likely due to their association with RCOR1 (PubMed:33417871). It also targets RCOR3, MIER2 and MIER3 for proteasomal degradation as well as associated proteins ZNF217 and RREB1 with degradation being dependent on the presence of an ELM2 domain in the target proteins (PubMed:36997086). The BCR(ARMC5) complex mediates premature transcription termination of transcripts that are unfavorably configured for transcriptional elongation by mediating ubiquitination of Pol II subunit POLR2A (PubMed:35687106, PubMed:38225631, PubMed:39504960, PubMed:39667934). Required for 'Lys-63'-linked ubiquitination of large ribosomal subunit protein MRPL12 (PubMed:37526061). |
| Cellular Location | Nucleus. Golgi apparatus. Cell projection, cilium, flagellum. Cytoplasm, cytoskeleton, spindle. Cytoplasm. Cytoplasm, cytoskeleton, microtubule organizing center, centrosome. Cytoplasm, cytoskeleton, spindle pole. Note=Detected along the length of the sperm flagellum and in the cytoplasm of the germ cells (PubMed:28395323). Predominantly found in the nucleus in interphase cells, found at the centrosome at late G2 or prophase, starts accumulating at the spindle poles in prometaphase and stays on the spindle poles and the mitotic spindle at metaphase (PubMed:23213400) |
| Tissue Location | Brain, spermatozoa, and testis (at protein level). Widely expressed. |

Thousands of laboratories across the world have published research that depended on the performance of antibodies from Abcepta to advance their research. Check out links to articles that cite our products in major peer-reviewed journals, organized by research category.
info@abcepta.com, and receive a free "I Love Antibodies" mug.
Provided below are standard protocols that you may find useful for product applications.
Background
CUL3 is a component of a ubiquitin E3 ligase that is essential for mitotic division (Sumara et al., 2007 [PubMed 17543862]).
References
Maerki, S., et al. J. Cell Biol. 187(6):791-800(2009)
Zhuang, M., et al. Mol. Cell 36(1):39-50(2009)
Chen, Y., et al. Mol. Cell 35(6):841-855(2009)
Eggler, A.L., et al. Biochem. J. 422(1):171-180(2009)
Jin, Z., et al. Cell 137(4):721-735(2009)
Beausoleil, S.A., et al. Proc. Natl. Acad. Sci. U.S.A. 101(33):12130-12135(2004)
Singer, J.D., et al. Genes Dev. 13(18):2375-2387(1999)
Ohta, T., et al. Mol. Cell 3(4):535-541(1999)
Du, M., et al. J. Biol. Chem. 273(38):24289-24292(1998)
Michel, J.J., et al. Cell Growth Differ. 9(6):435-449(1998)
Kipreos, E.T., et al. Cell 85(6):829-839(1996)
If you have used an Abcepta product and would like to share how it has performed, please click on the "Submit Review" button and provide the requested information. Our staff will examine and post your review and contact you if needed.
If you have any additional inquiries please email technical services at tech@abcepta.com.
Ordering Information
Other Products
Shipping Information
